


|
|
|
| Home | Initium | Angiogenesis | FAQ |
| TEM domain structure |
|---|
|
|
|
|
| TEM Sequence Analysis |
|---|
| name | from | to | source/E | description |
|---|---|---|---|---|
| Signalp | 1 | 27 | SignalP | |
| LRRNT | 29 | 72 | Pfam, E= 44 | The LLRNT domain is a cystein-rich domain that is found to border LRR-repeats. Due to the cystein conservation the hit is considered despite of the unreliable scoring. |
| LRR | 75 | 99 | PfamFrag, E=0.69 | LRRs are usually involved in protein-protein interactions. |
| LRR | 100 | 123 | Pfam, E=309e+02 | LRRs are usually involved in protein-protein interactions and cellular adhesion. |
| LRR | 124 | 147 | Pfam, E=0.0037 | LRRs are usually involved in protein-protein interactions and cellular adhesion. |
| LRR | 148 | 171 | Pfam, E=0.032 | LRRs are usually involved in protein-protein interactions and cellular adhesion. |
| LRRCT | 183 | 233 | Pfam, E=1.9e-08 | The LLRCT domain is a cystein-rich domain that is found to border LRR-repeats. |
| ig | 246 | 339 | PfamFrag, E=0.77 SMART, E=3.49e-03 |
Immunoglobulin-like domains may be involved in protein-protein and protein-ligand interactions. |
| HormR | 342 | 419 | SMART, E=4.57e-02 | The HormR domain is found in peptide hormone receptors and is a proposed ligand binding domain. |
| unknown | 419 | 510 | predator | The region is highly helical (>85 %) and contains a apolar span |
| unknown | 510 | 699 | predator | The region is predicted to be mostly unstructured (71 % coil), contains a LCR(P) from 556 to 580. Two apolar regions are predicted by toppred2 to be certain TM spans. The prediction of TM regions is not considered due to missing helix predictions in this region, the fact that the following GPS domain is supposed to be extracellular in proteins with various membrane topology (1-, 7-, 10TM), and due to known topology of proteins of the secretin family homologous in this region (PsiBlast, TEM9: HormR-GPS-7tm_2). |
| GPS | 699 | 751 | PFAM, E = 0.004 | The GPS domain can be found N-terminally of the 7 tm region in various secretin receptors. The domain in TEM9 contains one of the two putative di-S bridges found in most of the GPS domains. Functionally it could be a ligand binding and/or cleavage domain based on its similarity to G-protein-coupled receptor proteolytic site domain. |
| 7tm_2 | 762 | 1069 | PFAMsanger, E = 0.01 | The 7tm region defines TEM9 as a secretin family proteins. Characteristic residues were found to be conserved in TM segments, with the exception of TM6 (eg GxP motif in TM4). Most characterized receptors of the secretin family are coupled to G-proteins which activate both adenylyl cyclase and the phosphatidylinositol-calcium pathway. |
| cytoplasmic | 1069 | 1331 | The cytoplasmic sequence contains two vicinal cysteins typical for the GPCR-palmitoylation site. It is longer than in other G-protein-coupled receptors and shows no significant homologies. The composition is notable, as the region is highly LCR -rich: proline-, alanine-rich, and LCR with a positively charged cluster. Highly proline-rich regions usually serve as binding sites for Src homology (SH3) domains and other protein modules. A class II consensus-SH3 binding motif (XPpXPXR, p proline-preferred, X non-conserved residue) can be found (PHAPPR). | |
| 1331 |
| Remarks |
|---|
RESIDUE 1-339 LIGAND-BINDING DOMAIN:
RESIDUE 342-1069 LIGAND-BINDING AND RECEPTOR REGULATION BY SIMILARITY: The region is composed of HormR-linker-GPS-7tm_2.
GPS AS A CLEAVAGE SITE: The cleavage site in the GPS domain has been alocated in latrophilin CL-1,with the region around the putative cleavage site (HLGN) being most highly conserved. Several proteins containing this cystein-rich region have been shown to be cleaved :CL-1, 2, 3, CD97, making it likely that TEM9 could be processed as well. The subunits resulting from the CL-1 cleavage have been shown to remain associated. From the four conserved cystein residues mostly found in GPS domains only two cysteins are conserved in Tem9, human PKD1 and in C.elegans F31D5.4, F31D5.5.
| Sequence Information |
|---|
| TEM | TEM9 | ||||||
Reliability of Gen2Tag mapping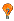 |
|
||||||
| Analysis |
|
||||||
| Protein Description |
|
||||||
| Protein Sequence |
MRGAPARLLLPLLPWLLLLLAPEARGAPGCPLSIRSCKCSGERPKGLSGGVPGPARRRVVCSGGDLPEPP EPGLLPNGTVTLLLSNNKITGLRNGSFLGLSLLEKLDLRNNIISTVQPGAFLGLGELKRLDLSNNRIGCL TSETFQGLPRLLRLNISGNIFSSLQPGVFDELPALKVVDLGTEFLTCDCHLRWLLPWAQNRSLQLSEHTL CAYPSALHAQALGSLQEAQLCCEGALELHTHHLIPSLRQVVFQGDRLPFQCSASYLGNDTRIRWYHNRAP VEGDEQAGILLAESLIHDCTFITSELTLSHIGVWASGEWECTVSMAQGNASKKVEIVVLETSASYCPAER VANNRGDFRWPRTLAGITAYQSCLQYPFTSVPLGGGAPGTRASRRCDRAGRWEPGDYSHCLYTNDITRVL YTFVLMPINASNALTLAHQLRVYTAEAASFSDMMDVVYVAQMIQKFLGYVDQIKELVEVMVDMASNLMLV DEHLLWLAQREDKACSRIVGALERIGGAALSPHAQHISVNARNVALEAYLIKPHSYVGLTCTAFQRREGG VPGTRPGSPGQNPPPEPEPPADQQLRFRCTTGRPNVSLSSFHIKNSVALASIQLPPSLFSSLPAALAPPV PPDCTLQLLVFRNGRLFHSHSNTSRPGAAGPGKRRGVATPVIFAGTSGCGVGNLTEPVAVSLRHWAEGAE PVAAWWSQEGPGEAGGWTSEGCQLRSSQPNVSALHCQHLGNVAVLMELSAFPREVGGAGAGLHPVVYPCT ALLLLCLFATIITYILNHSSIRVSRKGWHMLLNLCFHIAMTSAVFAGGITLTNYQMVCQAVGITLHYSSL STLLWMGVKARVLHKELTWRAPPPQEGDPALPTPSPMLRFYLIAGGIPLIICGITAAVNIHNYRDHSPYC WLVWRPSLGAFYIPVALILLITWIYFLCAGLRLRGPLAQNPKAGNSRASLEAGEELRGSTRLRGSGPLLS DSGSLLATGSARVGTPGPPEDGDSLYSPGVQLGALVTTHFLYLAMWACGALAVSQRWLPRVVCSCLYGVA ASALGLFVFTHHCARRRDVRASWRACCPPASPAAPHAPPRALPAAAEDGSPVFGEGPPSLKSSPSGSSGH PLALGPCKLTNLQLAQSQVCEAGAAAGGEGEPEPAGTRGNLAHRHPNNVHHGRRAHKSRAKGHRAGEACG KNRLKALRGGAAGALELLSSESGSLHNSPTDSYLGSSRNSPGAGLQLEGEPMLTPSEGSDTSAAPLSEAG RAGQRRSASRDSLKGGGALEKESHRRSYPLNAASLNGAPKGGKYDDVTLMGAEVASGGCMKTGLWKSETT V |
||||||
|
SAGE-Tag |
CATG-TTTTTAAGAAC | ||||||
| EST/cluster Description |
|
||||||
| EST/cluster Sequence |
ACCACCTCATCCCGTCCCTACGCCAAGTGGTGTTCCAGGGGGATCGGCTGCCCTTCCAGTGCTCTGCCAG CTACCTGGGCAACGACACCCGCATCCGCTGGTACCACAACCGAGCCCCTGTGGAGGGTGATGAGCAGGCG GGCATCCTCCTGGCCGAGAGCCTCATCCACGACTGCACCTTCATCACCAGTGAGCTGACGCTGTCTCACA TCGGCGTGTGGGCCTCAGGCGAGTGGGAGTGCACCGTGTCCATGGCCCAAGGCAACGCCAGCAAGAAGGT GGAGATCGTGGTGCTGGAGACCTCTGCCTCCTACTGCCCCGCCGAGCGTGTTGCCAACAACCGCGGGGAC TTCAGGTGGCCCCGAACTCTGGCTGGCATCACAGCCTACCAGTCCTGCCTGCAGTATCCCTTCACCTCAG TGCCCCTGGGCGGGGGTGCCCCGGGCACCCGAGCCTCCCGCCGGTGTGACCGTGCCGGCCGCTGGGAGCC AGGGGACTACTCCCACTGTCTCTACACCAACGACATCACCAGGGTGCTGTACACCTTCGTGCTGATGCCC ATCAATGCCTCCAATGCGCTGACCCTGGCTCACCAGCTGCGCGTGTACACAGCCGAGGCCGCTAGCTTTT CAGACATGATGGATGTAGTCTATGTGGCTCAGATGATCCAGAAATTTTTGGGTTATGTCGACCAGATCAA AGAGCTGGTAGAGGTGATGGTGGACATGGCCAGCAACCTGATGCTGGTGGACGAGCACCTGCTGTGGCTG GCCCAGCGCGAGGACAAGGCCTGCAGCCGCATCGTGGGTGCCCTGGAGCGCATTGGGGGGGCCGCCCTCA GCCCCCATGCCCAGCACATCTCAGTGAATGCGAGGAACGTGGCATTGGAGGCCTACCTCATCAAGCCGCA CAGCTACGTGGGCCTGACCTGCACAGCCTTCCAGAGGAGGGAGGGAGGGGTGCCGGGCACACGGCCAGGA AGCCCTGGCCAGAACCCCCCACCTGAGCCCGAGCCCCCAGCTGACCAGCAGCTCCGCTTCCGCTGCACCA CCGGGAGGCCCAATGTTTCTCTGTCGTCCTTCCACATCAAGAACAGCGTGGCCCTGGCCTCCATCCAGCT GCCCCCGAGTCTATTCTCATCCCTTCCGGCTGCCCTGGCTCCCCCGGTGCCCCCAGACTGCACCCTGCAA CTGCTCGTCTTCCGAAATGGCCGCCTCTTCCACAGCCACAGCAACACCTCCCGCCCTGGAGCTGCTGGGC CTGGCAAGAGGCGTGGCGTGGCCACCCCCGTCATCTTCGCAGGAACCAGTGGCTGTGGCGTGGGAAACCT GACAGAGCCAGTGGCCGTTTCGCTGCGGCACTGGGCTGAGGGAGCCGAACCTGTGGCCGCTTGGTGGAGC CAGGAGGGGCCCGGGGAGGCTGGGGGCTGGACCTCGGAGGGCTGCCAGCTCCGCTCCAGCCAGCCCAATG TCAGCGCCCTGCACTGCCAGCACTTGGGCAATGTGGCCGTGCTCATGGAGCTGAGCGCCTTTCCCAGGGA GGTGGGGGGCGCCGGGGCAGGGCTGCACCCCGTGGTATACCCCTGCACGGCCTTGCTGCTGCTCTGCCTC TTCGCCACCATCATCACCTACATCCTCAACCACAGCTCCATCCGTGTGTCCCGGAAAGGCTGGCACATGC TGCTGAACTTGTGCTTCCACATAGCCATGACCTCTGCTGTCTTTGCGGGGGGCATCACACTCACCAACTA CCAGATGGTCTGCCAGGCGGTGGGCATCACCCTGCACTACTCCTCCCTATCCACGCTGCTCTGGATGGGC GTGAAGGCGCGAGTGCTCCATAAGGAGCTCACCTGGAGGGCACCCCCTCCGCAAGAAGGGGACCCCGCTC TGCCTACTCCCAGTCCTATGCTCCGCTGCTGGCTGGTGTGGCGTCCAAGCCTTGGCGCCTTCTACATCCC TGTGGCTTTGATTCTGCTCATCACCTGGATCTATTTCCTGTGCGCCGGGCTACGCTTACGGGGTCCTCTG GCACAGAACCCCAAGGCGGGCAACAGCAGGGCCTCCCTGGAGGCAGGGGAGGAGCTGAGGGGTTCCACCA GGCTCAGGGGCAGCGGCCCCCTCCTGAGTGACTCAGGTTCCCTTCTTGCTACTGGGAGCGCGCGAGTGGG GACGCCCGGGCCCCCGGAGGATGGTGACAGCCTCTATTCTCCGGGAGTCCAGCTAGGGGCGCTGGTGACC ACGCACTTCCTGTACTTGGCCATGTGGGCCTGCGGGGCTCTGGCAGTGTCCCAGCGCTGGCTGCCCCGGG TGGTGTGCAGCTGCTTGTACGGGGTGGCAGCCTCCGCCCTGGGCCTCTTCGTCTTCACTCACCACTGTGC CAGGCGGAGGGACGTGAGAGCCTCGTGGCGCGCCTGCTGCCCCCCTGCCTCTCCCGCGGCCCCCCATGCC CCGCCCCGGGCCCTGCCCGCCGCCGCAGAGGACGGTTCCCCGGTGTTCGGGGAGGGGCCCCCCTCCCTCA AGTCCTCCCCAAGCGGCAGCAGCGGCCATCCGCTGGCTCTGGGCCCCTGCAAGCTCACCAACCTGCAGCT GGCCCAGAGTCAGGTGTGCGAGGCGGGGGCGGCGGCCGGCGGGGAAGGAGAGCCGGAGCCGGCGGGCACC CGGGGAAACCTCGCCCACCGCCACCCCAACAACGTGCACCACGGGCGTCGGGCGCACAAGAGCCGGGCCA AGGGACACCGCGCGGGGGAGGCCTGCGGCAAGAACCGGCTCAAGGCCCTGCGCGGGGGCGCGGCGGGGGC GCTGGAGCTGCTGTCCAGCGAGAGCGGTAGTCTGCACAACAGCCCCACCGACAGCTACCTGGGCAGCAGC CGCAACAGCCCGGGCGCCGGCCTGCAGCTGGAAGGCGAGCCCATGCTCACGCCGTCCGAGGGCAGCGACA CCAGCGCCGCGCCGCTTTCTGAGGCGGGCCGGGCAGGCCAGCGCCGCAGCGCCAGCCGCGACAGTCTCAA GGGCGGCGGCGCGCTGGAGAAGGAGAGCCATCGCCGCTCGTACCCGCTCAACGCCGCCAGCCTAAACGGC GCCCCCAAGGGGGGCAAGTACGACGACGTCACCCTGATGGGCGCGGAGGTAGCCAGCGGCGGCTGCATGA AGACCGGACTCTGGAAGAGCGAAACTACCGTCTAAGGTGGGGCGGGCGACGCGGTAGACGGGCTGGCCAC GCGGCTCGTTCCCCCGCTCCTCGGGGCCCTCCAAGGTGTCTCCGTAGTCAGCAGGTTGGAGGCAGAGGAG CCGATGGCTGGAGGAAGCCCACAGGCGGATGTTCCCCACTTGCCTAGAGGGCATCCCTCTGGGGTAGCGA CAGACAATCCCAGAAACACGCATAATACATTTCCGTCCAGCCCGGGGCAGTCTGACTGTCGGTGCCCTCC CAGGAACGGGGAAGGCCTCCGTCTGTGTGAAAGGGCACAGCACATCCCAGGTGCACCCTCCCCAAGTACT CCCACCCCGCCTACTGTCCATGCGGCCTCACTGGGGGCCATCAGCCTCACCAGCAAAGCAGAGATGAGAG CGTGGGAACTGTGTTCTTTCCTCCCTGCCCTCTACTGATTTCAGCCCAGCCCCTGCCTAGATCCTAGGTC CCTTTTCCTCCCGAGTTTGGCTGGCACGAGAGCTAGCCCAGCACATGAAGCAGGTGATGTTAAGTCACAA GGTGCTGCTTTTCAGATCCACTATGCAAGAGGGGAGGGTGGGGCCACGTGAAAGGCAGCTCTAGACATCA ACCAGTCCTGGGGGAGGGGAGTGGGAACCGGGCACAACTAGGAACAATGCCACCATTCCCACAGGAGTGG TACTTAAACCAGACAGCAGGGTTCAGAGGTGGCACACCGGGACAAAGCTGAGGCCCTGCACCTCAACAGC TGACTGCCAGGTGCCTGTGGGTGAACTGAGGGGAGTAGAGGGAGAGGGCAGGTGGAACTGGGGCAGAATC TAGTCATGCCCTAAAGCTAGTCCTGTAAACAATGGTGCCCCAGAAAGCTGCAGGTGGTGTTTGGAGAAGC AGTTACTTTTCAGTTACAAGACCCATCTCCCTAGTCTCAGCCTTACAACACCACGGGACTAAGGAAGAGC ACTTCCTTGCCTCCGTAAGGCCAGAGGAAGAACCATCCCAATCATTTGATCTCCAGCTCCACAGTAGAGA GAAACCTACAAAATGTCAAACCAGCTTCCCGACTCCCAGGAGCTCAAGCCAAGCCCAGAGGCAGTGGCTG GGGTCCCTGCAGGTCATGAGGGGCCTATGCCTTTACTCCTTTTAAACACCAGCACCCGTCTTTTCCCCAA CCTAAAACCAACCACCAGCATTTCACTACAGGACCAAATGGAAACCGAGGGAACCCTGGGTCTTGGGAAG AACAACAGGAAACCAAGGTCTGACCTAGGGTTCCCTCCCAGTCTTCACATCACTCTGGCCTCATCACCAA GGTGACAGAGGACACAGGGGAGGGGGAAAACCCACACACACTCCTTGGAATGGGTCCTGTTATTTATGCT TGCTGCACAGACATATTAGAAGAAAAAAAAAAGCTTTGTATTATTCTTCCACATATGCTGGCTGCTGTTT ACACACCCTGCCAATGCCTTAGCACTGGAGAGCTTTTTGCAATATGCTGGGGAAAGGGGAGGGAGGGAAT GAAAGTGCCAAAGAAAACATGTTTTTAAGAACTCGGGTTTTATACAATAGAATGTTTTCTAGCAGATGCC TCTTGTTTTAATATATTAAAATTTTGCAAAGCCCTTTG |
||||||
| Genomic Sequence Description |
|
||||||
| Genomic Sequence |
|
| Literature |
|---|
| PubMed:99121098 | Structural requirements for alpha-latrotoxin binding and alpha-latrotoxin-stimulated secretion. A study with calcium-independent receptor of alpha-latrotoxin (CIRL) deletion mutants. Krasnoperov V, Bittner MA, Holz RW, Chepurny O, Petrenko AG. J Biol Chem 1999 Feb 5;274(6):3590-6 |
| PubMed:9904765 | alpha-Latrotoxin receptor CIRL/latrophilin 1 (CL1) defines an unusual family of ubiquitous G-protein-linked receptors. G-protein coupling not required for triggering exocytosis. Sugita S, Ichtchenko K, Khvotchev M, Sudhof TC. J Biol Chem 1998 Dec 4;273(49):32715-24 |
| PubMed:10469603 | A latrophilin/CL-1-like GPS domain in polycystin-1. Ponting CP, Hofmann K, Bork P. Curr Biol. 1999 Aug 26;9(16):R585-8. |
| PubMed:11086986 | Control of dendritic field formation in Drosophila: the roles of flamingo and competition between homologous neurons. Gao FB, Kohwi M, Brenman JE, Jan LY, Jan YN. Neuron. 2000 Oct;28(1):91-101. |
| PubMed:10556066 | The Drosophila tissue polarity gene starry night encodes a member of the protocadherin family. Chae J, Kim MJ, Goo JH, Collier S, Gubb D, Charlton J, Adler PN, Park WJ. Development. 1999 Dec;126(23):5421-9. |
| PubMed:20202599 | Large exons encoding multiple ectodomains are a characteristic feature of protocadherin genes. Wu Q, Maniatis T Proc Natl Acad Sci U S A 2000 Mar 28;97(7):3124-9 |
| PubMed:7526465 | Two binding orientations for peptides to the Src SH3 domain: development of a general model for SH3-ligand interactions. Feng S, Chen JK, Yu H, Simon JA, Schreiber SL. Science. 1994 Nov 18;266(5188):1241-7. |
| Home | Initium | Angiogenesis | FAQ |

{kind=link}