|
|
|
| Home | Initium | Angiogenesis | FAQ |
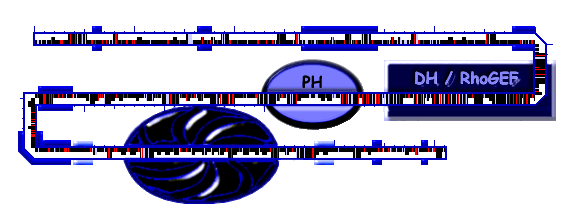
| TEM domain structure |
|---|
|
|
|
|
| TEM Sequence Analysis |
|---|
| name | from | to | source/E | description |
|---|---|---|---|---|
| unknown | 1 | 517 | BLASTP without filtering | No known domain matches this region. It is LCR-rich (P) and 8 potential SH3 binding motif sequences (PXXP) cluster in the region. A potential 14-3-3-binding motif (RSXSXP) is found at position 179-184. The actin-binding protein profilin binds polyproline rich sequence tracts containing two or more ZPPX motifs in a stretch of no more than 100 residues (Z- P G A or charged, X- preferentially hydrophobic). The N-terminal of TEM5old4 contains 7 ZPPX motifs (64, 102, 343, 422, 428, 483, 490), 4 with a hydrophobic amino-acid at motif-position 4 (64, 102, 342, 421, 427). The function of the proline-rich N-terminal region remains unknown, although a partially sequence-similar proline-rich region in FGD1 was shown to mediate subcellular localization to the actin cytoskeleton. |
| 9 | 20 | SEG | LCR(SL) | |
| 30 | 55 | predator | HELIX | |
| 57 | 68 | SEG | LCR(DPG) | |
| 162 | 176 | SEG | LCR(GS) | |
| 277 | 344 | SEG25 | LCR(PSGE) | |
| 478 | 502 | SEG | LCR(PA) | |
| DH/RhoGEF | 517 | 700 | PFAM, E=7.5e-43 |
This region contains a potential GEF-domain for Rho/Rac/Cdc42-like GTPases. DH domains are found to function not only as GEFs, but additionally to recruit GTPases to targets and locations. The DH-domain of TEM5 shows closest homology to bcr, ect2, pbl (the latter two are Rho-specific). |
| PH | 718 | 930 | IMPALA1.1, E=3.3 (722-822) | The hit is not reliable, but as PH domains are invariably found C-terminally to DH domains it can be considered. The assignment is based additionaly on a Psi-Blast search of the region 700-1000 with the inclusion condition E=0.002 collecting the PH domain of human bcr. While the DH domain is responsible for catalytic activity, the PH domain has proposed functions in proper localization or full catalytic activity of the DH domain. |
| unknown | 930 | 1012 | 44% identity in CG7397 | This region of unknown function contains a motif of potential significance due to its 100% preserved sequence compared to CG7397: VWVCNSDGYVGQVC |
| 1012 | 1048 | SEG | No known domain matches this region. It is LCR enriched in proline and alanine. | |
| 1083 | 1134 | SEG | No known domain matches this region. It is LCR enriched in serine and contains a predicted high scoring PEST domain. | |
| apolar region | 1135 | 1155 | TOPPREDM(1.22) | The following 7bladedbeta-propeler domain seems to be bordered by two apolar, unstructured regions. |
| 7 bladed beta-propeler | 1191 | 1362 | IMPALA-SCOP, E= 1e-06 | The region is predicted to form a 7 bladed beta-propeler, which is a structure frequently involved in multiple protein-interaction formation. The secondary structures adopted in this region can be described are predicted as follows: alpha-helix (1251-1239) & 7predicted beta-str (1239-1362). The region shows homology to the last 400 aa of JNK/SAPK-associated protein-1 (see below). |
| apolar region | 1380 | 1394 | TOPPREDM(1.539) | The preceeding 7bladedbeta-propeler domain seems to be bordered by two apolar, unstructured regions. |
| unknown | 1394 | 1493 | blastp | This region shows homology to CG7397 and JNK/SAPK-associated protein-1, the later of the two being more indicative on its potential function as it is studied. Seven potential SH3 binding motif sequences (PXXP) cluster in this region. |
| 1510 |
| Remarks |
|---|
The DH domain is known to confer catalytic GEF-activity. Family members show great variation in target-specificity: with either a broad GE-specificity (Dbl, Ect2), or a restricted GE-specificity (FGD1-cdc 42 specific). An additional function of DH domains besides the catalytic activity, could be the recruitment of GTPases to targets and locations. Closest homology of studied proteins to the TEM5 DH domain can be found in bcr(AAB60390.1), pbl[Drosophila](AAF50508.1), ect2(NP_031926.1) with identities in the range of 30%.
The C-terminal beta-propeller forming region shows 27% identity to the C-terminal region in JSAP1 (BAB16676.1) and mSYD2 (AF262046).
A proline, serine rich LCR, which lies inbetween the DH/PH and the beta-propeller forming region, shows significantly scoring PEST sites in the region 1019 to 1158, the best being 1076 to 1104 (+14.31).
Due to the potential integrative function of TEM5 in signaling and the fact that the proline-rich regions of Tem5 contain several minimal SH3 binding site (PXXP) the protein associates with Src homology 3-containing signaling proteins. Additionally, an match with the exact binding site for 14-3-3 within the aminoterminal region of Tem5 might indicate a role for 14-3-3 proteins in regulation of the protein activity.
| Sequence Information |
|---|
| TEM | TEM5 | ||||||
Reliability of Gen2Tag mapping |
|
||||||
| Analysis |
|
||||||
| Protein Description |
|
||||||
| Protein Sequence |
MARRLPRTSALKSSSSELLLTGPGAEEDPLPLIVQDQYVQEARQVFEKIQRMGAQQDDGSDAPPGSPDWA GDVTRGQRSQEELSGPESSLTDEGIGADPEPPVAAFCGLGTTGMWRPLSSSSAQTNHHGPGTEDSLGGWA LVSPETPPTPGALRRRRKVPPSGSGGSEFSNGEAGEAYRSLSDPIPQRHRAATSEEPTGFSVDSNLLGSL SPKTGLPATSAMDEGLTSGHSDWSVGSEESKGYQEVIQSIVQGPGTLGRVVDDRIAGKAPKKKSLSDPSR RGELAGPGFEGPGGEPIREVEPMLPPSSSEPILVEQRAEPEEPGATRSRAQSERALPEALPPPATAHRNF HLDPKLADILSPRLIRRGSKKRPARSSHQELRRDEGSQDQTGSLSRARPSSRHVRHASVPATFMPIVVPE PPTSVGPPVAVPEPIGFPTRAHPTLQAPSLEDVTKQYMLNLHSGEVPAPVPVDMPCLPLAAPPSAEAKPP EAARPADEPTPASKCCSKPQVDMRKHVAMTLLDTEQSYVESLRTLMQGYMQPLKQPENSVLCDPSLVDEI FDQIPELLEHHEQFLEQVRHCMQTWHAQQKVGALLVQSFSKDVLVNIYSAYIDNFLNAKDAVRVAKEARP AFLKFLEQSMRENKEKQALSDLMIKPVQRIPRYELLVKDLLKHTPEDHPDHPLLLEAQRNIKQVAERINK GVRSAEEAERHARVLQEIEAHIEGMEDLQAPLRRFLRQEMVIEVKAIGGKKDRSLFLFTDLIVCTTLKRK SGSLRRSSMSLYTAASVIDTASKYKMLWKLPLEDADIIKGASQATNRENIQKAISRLDEDLTTLGQMSKL SESLGFPHQSLDDALRDLSAAMHRDLSEKQALCYALSFPPTKLELCATRPEGTDSYIFEFPHPDARLGFE QAFDEAKRKLASSKSCLDPEFLKAIPIMKTRSGMQFSCAAPTLNSCPEPSPEVWVCNSDGYVGQVCLLSL RAEPDVEACIAVCSARILCIGAVPGLQPRCHREPPPSLRSPPETAPEPAGPELDVEAAADEEAATLAEPG PQPCLHISIAGSGLEMTPGLGEGDPRPELVPFDSDSDDESSPSPSGTLQSQASRSTISSSFGNEETPSSK EATAETTSSEEEQEPGFLPLSGSFGPGGPCGTSPMDGRALRRSSHGSFTRGSLEDLLSVDPEAYQSSVWL GTEDGCVHVYQSSDSIRDRRNSMKLQHAASVTCILYLNNQVFVSLANGELVVYQREAGHFWDPQNFKSVT LGTQGSPITKMVSVGGRLWCGCQNRVLVLSPDTLQLEHMFYVGQDSSRCVACMVDSSLGVWVTLKGSAHV CLYHPDTFEQLAEVDVTPPVHRMLAGSDAIIRQHKAACLRITALLVCEELLWVGTSAGVVLTMPTSPGTV SCPRAPLSPTGLGQGHTGHVRFLAAVQLPDGFNLLCPTPPPPPDTGPEKLPSLEHRDSPWHRGPAPARPK MLVISGGDGYEDFRLSSGGGSSSETVGRDDSTNHLLLWRV |
||||||
|
SAGE-Tag |
CATG-TATTAACTCTC | ||||||
| EST/cluster Description |
|
||||||
| EST/cluster Sequence |
ACGACCTATGGTCTAGTAGGGGTTCTGGGGGCTGGGGCGTGTACCGCTCCCCTAGCTTTGGAGCTGGGGA AGGGCTCCTGCGGTCCCAGGCTCGAACCCGTGCCAAAGGACCTGGAGGCACCTCTAGGGCATTGAGGGAT GGAGGATTTGAGCCTGAAAGAGTCGACAGCGGAAGTCCCTGTCAAATCCAGATATCGCCTCAGAGACCCT GACGCTTCTCAGTTTCCTGCGCTCAGACCTTTCAGAGCTGAGGGTCCGAAAACCTGGTGGGAGCTCCGGG GACCGTGGAAGCAACCCCCTAGATGGCAGAGACTCACCATCCGCAGGTGGCCCTGTGGGGCAACTTGAAC CCATACCCATCCCAGCCCCAGCATCACCTGGCACGCGCCCCACACTCAAGGACTTGACAGCCACTCTGCG GAGAGCAAAGTCATTCACCTGCTCTGAGAAGCCCATGGCCCGCCGCCTGCCCCGCACCAGTGCTCTGAAG TCCAGCTCCTCCGAGCTCCTGCTCACAGGCCCTGGTGCCGAGGAGGATCCGCTGCCCCTCATCGTCCAGG ACCAATATGTGCAGGAGGCCCGCCAGGTTTTTGAGAAGATCCAGCGCATGGGTGCCCAACAAGATGATGG AAGCGATGCCCCCCCTGGAAGCCCTGACTGGGCAGGGGATGTGACCCGAGGGCAGCGGTCCCAGGAGGAG CTCTCAGGCCCTGAGTCCAGTCTGACAGATGAAGGCATTGGGGCAGACCCTGAGCCTCCTGTTGCAGCAT TTTGCGGCCTGGGTACCACAGGGATGTGGCGACCTCTTTCCTCATCCTCGGCCCAGACGAACCACCATGG CCCTGGGACTGAGGACAGTCTGGGCGGGTGGGCCCTGGTGTCGCCTGAGACCCCTCCCACACCAGGTGCC CTCCGCCGACGACGCAAAGTCCCACCTTCAGGTTCTGGTGGGAGCGAATTTAGCAATGGGGAGGCAGGGG AGGCCTACAGGTCCCTGAGTGACCCAATTCCTCAGCGCCACCGGGCTGCCACCTCTGAAGAGCCTACTGG GTTCTCTGTGGACAGCAACCTCCTGGGCTCACTGAGCCCCAAGACAGGGCTCCCTGCCACCTCAGCCATG GATGAGGGCTTGACCAGTGGTCACAGTGACTGGTCTGTGGGCAGTGAAGAGAGCAAGGGATATCAGGAGG TTATTCAGAGCATAGTTCAGGGGCCTGGCACCCTGGGGCGTGTGGTGGACGACAGGATTGCTGGCAAAGC CCCCAAGAAGAAATCCCTGAGTGACCCCAGCCGCCGTGGGGAGCTGGCTGGGCCTGGATTCGAGGGCCCT GGAGGGGAGCCCATCCGAGAAGTTGAGCCCATGCTGCCTCCATCCAGCAGCGAGCCCATCCTTGTAGAGC AGCGGGCAGAGCCAGAAGAACCTGGTGCCACCAGGAGCCGGGCACAGTCTGAAAGGGCCCTACCTGAGGC TCTGCCTCCCCCTGCCACTGCCCACCGAAACTTTCACCTTGACCCCAAGCTGGCTGACATTCTGTCCCCG AGGCTAATCCGCCGAGGCTCCAAGAAGCGCCCAGCTCGGAGTAGTCACCAGGAGCTTCGGAGAGACGAGG GCAGTCAGGACCAGACTGGCAGCCTGTCTCGGGCCCGGCCCTCCTCCAGACACGTTCGCCATGCCAGTGT GCCCGCCACATTTATGCCTATTGTGGTGCCTGAGCCACCAACTTCTGTTGGTCCCCCTGTGGCTGTGCCA GAACCCATAGGCTTCCCTACCCGAGCCCATCCCACGTTGCAGGCACCATCGCTCGAGGACGTCACCAAGC AGTACATGCTGAACCTGCACTCCGGTGAGGTCCCTGCCCCAGTGCCAGTGGACATGCCCTGCTTGCCTCT GGCTGCACCGCCCTCTGCTGAGGCCAAGCCCCCTGAGGCAGCTCGGCCTGCAGATGAGCCTACCCCTGCC AGCAAGTGCTGCAGCAAGCCACAGGTGGACATGCGGAAGCACGTGGCCATGACCCTGCTGGACACAGAGC AGTCGTATGTGGAGTCGCTGCGCACCCTGATGCAGGGCTACATGCAGCCGCTGAAGCAGCCAGAGAACTC CGTGCTCTGTGACCCTTCACTGGTGGACGAGATCTTCGACCAGATCCCCGAGCTCCTGGAGCACCACGAG CAATTCCTGGAGCAGGTTCGGCACTGCATGCAGACCTGGCATGCCCAGCAGAAGGTGGGAGCCCTGCTCG TCCAGTCGTTCTCCAAGGATGTCCTAGTAAACATCTATTCTGCCTATATCGATAACTTCCTCAATGCAAA GGATGCTGTGCGTGTGGCCAAGGAGGCGAGGCCTGCCTTTCTCAAGTTCCTAGAGCAAAGCATGCGTGAG AACAAGGAGAAGCAGGCGCTGTCTGACCTCATGATCAAGCCTGTGCAGCGGATCCCACGCTACGAGCTTC TGGTGAAGGACCTCCTGAAGCATACACCTGAGGACCACCCGGACCATCCACTCCTGCTGGAGGCGCAGCG GAACATCAAGCAGGTGGCTGAGCGCATCAACAAGGGTGTGCGGAGTGCCGAGGAGGCGGAGCGCCATGCC CGTGTGCTGCAGGAGATAGAGGCTCACATCGAGGGCATGGAGGATCTCCAGGCCCCTCTGCGGCGGTTCC TGAGACAGGAGATGGTCATTGAAGTGAAGGCGATCGGTGGCAAGAAGGACCGGTCTCTCTTCCTGTTCAC GGACCTCATCGTCTGCACCACTCTGAAGCGAAAGTCAGGCTCCCTGCGGCGCAGCTCCATGAGCCTGTAC ACGGCAGCCAGTGTCATTGACACAGCCAGCAAGTACAAGATGCTGTGGAAGCTGCCGCTGGAAGACGCAG ACATCATCAAAGGGGCATCCCAAGCCACCAATCGGGAGAACATCCAGAAGGCCATCAGCCGCCTTGATGA GGACCTCACCACCCTGGGCCAAATGAGCAAGCTCTCTGAGAGCCTTGGTTTCCCCCACCAGAGCCTGGAC GATGCACTGCGGGACCTCTCAGCTGCCATGCACCGGGACCTGTCGGAGAAGCAGGCGCTGTGCTACGCGC TTTCCTTCCCGCCAACCAAGCTGGAGCTGTGCGCCACTCGGCCCGAGGGCACCGACTCCTACATTTTTGA GTTCCCTCACCCTGACGCCCGCCTTGGTTTTGAACAGGCCTTCGATGAGGCCAAGAGGAAGCTGGCATCC AGCAAAAGCTGTCTAGACCCTGAGTTCCTGAAGGCCATCCCCATCATGAAAACCCGCAGTGGCATGCAGT TCTCCTGTGCGGCTCCCACCCTGAACAGCTGCCCGGAGCCCTCGCCTGAGGTATGGGTCTGCAACAGCGA CGGCTACGTGGGCCAGGTGTGCCTGCTGAGCCTGCGCGCCGAGCCGGACGTGGAGGCCTGCATCGCCGTC TGTTCCGCCCGCATCCTCTGCATCGGGGCGGTGCCCGGGCTGCAGCCTCGCTGCCACCGGGAGCCTCCTC CGTCGCTGAGGAGTCCTCCAGAGACGGCACCGGAGCCCGCCGGGCCGGAGCTGGACGTCGAGGCCGCTGC AGACGAGGAAGCCGCGACGCTCGCGGAGCCGGGGCCGCAGCCCTGCCTTCACATCTCCATTGCAGGCTCG GGCTTGGAGATGACGCCGGGCCTCGGCGAGGGTGACCCCCGCCCAGAGCTGGTGCCCTTTGACAGTGACT CTGACGATGAGTCTTCGCCCAGCCCCTCGGGGACGCTGCAGAGCCAGGCCAGCCGGTCCACCATCTCCTC CAGCTTTGGCAATGAGGAGACCCCGAGTTCCAAGGAGGCCACGGCAGAGACCACCAGCTCAGAGGAGGAG CAGGAGCCAGGCTTCCTGCCACTGTCTGGCTCCTTTGGGCCTGGTGGTCCCTGCGGCACCAGCCCAATGG ATGGGAGAGCCCTTCGCCGCTCCAGCCACGGCTCCTTCACCCGGGGCAGCCTTGAGGACCTGCTGAGTGT CGACCCTGAGGCCTACCAGAGCTCCGTGTGGCTGGGCACTGAGGATGGCTGTGTCCACGTGTACCAGTCC TCCGACAGCATCCGTGACCGCAGGAACAGCATGAAGCTCCAGCATGCGGCCTCTGTGACCTGCATCTTGT ATCTGAATAACCAGGTGTTTGTGTCTCTGGCCAATGGAGAGCTTGTGGTCTACCAAAGGGAAGCAGGCCA TTTCTGGGACCCCCAGAACTTCAAATCAGTGACCTTGGGCACCCAGGGGAGCCCCATCACCAAGATGGTA TCTGTGGGTGGGCGGCTGTGGTGTGGCTGCCAGAACCGAGTCCTTGTCCTGAGCCCTGACACGCTGCAGC TGGAGCACATGTTTTACGTGGGTCAGGATTCAAGCCGCTGCGTGGCTTGCATGGTGGACTCCAGCCTGGG TGTGTGGGTGACATTGAAAGGTAGTGCCCACGTGTGTCTCTACCATCCAGACACCTTTGAGCAGCTGGCA GAAGTAGACGTCACTCCTCCCGTGCACAGGATGCTGGCAGGCTCGGATGCCATCATCCGGCAGCACAAGG CTGCCTGTCTGCGAATCACAGCGCTGCTGGTGTGTGAGGAGCTGCTGTGGGTGGGCACCAGTGCTGGTGT CGTCCTCACCATGCCCACTTCGCCCGGTACTGTCAGCTGCCCACGGGCACCACTCAGTCCCACAGGCCTC GGCCAGGGACACACCGGCCACGTCCGCTTCTTGGCTGCAGTCCAGCTGCCAGATGGCTTCAACCTGCTCT GCCCAACCCCACCACCTCCCCCAGACACAGGCCCCGAGAAGCTGCCATCACTGGAGCACCGGGACTCCCC TTGGCACCGAGGCCCCGCCCCTGCCAGGCCTAAAATGCTGGTTATCAGTGGAGGTGATGGCTATGAGGAC TTCCGACTCAGCAGTGGGGGCGGCAGCAGCAGTGAGACTGTGGGTCGAGACGACAGCACAAACCACCTCC TCCTGTGGAGGGTGTGACCCTGTCTGCCGTGGCCCAGGACTCGCCCGCCCACCTGCCTTCAGCCTGCTTG CCTCTCCCTAGCCCACACGCAGACTTTGACCAGGAGTATCCAGCCAGGGGCACACATGTGCCTGCGTGGG CTCTGCCTTGTCTTCGCGGAAGCATTCCTGATGGAACACCCACTGGCCAGCCAGGCCATGGCTTCTCCCG ACCCTCTGGCTGCCCCGGTGCTTCCAGTCATGATCGGGTGGGGGACATGTGGGCTGACCAGGACCTCTGA CCCTGGAGCTTCTACCAAAGACACAGCTGGGTCTGGACCCCACGGGGCTGGGGAGGGCCATGTGCAATAT TTGGAGGGTTTTCTGGAGGGCAGCAGGAAGGCTGGGGAATTCCCCATGTACAGTATTTATGTTTCTTTTT AGATGTGTACCTTCCCAAGCACTTATTTATGCAGTGACCTGGTCACCTGGGGTGGGGGTGATTTGAGGAA ATGACATGAGGAAAAGAAACCTATTCCTGCCCTGGGGACCACCCTGGGACTCTAACCAAGCCTTCCTGGA GGGACCCATGCGCCCCTGAGCCCCATTCCATTCATACAGACACACACGTACGCACACTGCATGTCCAAGG CCCTAAACATTGCCCGTTGACATAAACTTTCCAGGGCCCCAGCCTGATGGGGCTGCCCTCAGTCCTCTAG ATCAAGATGCTGACTATTAGGGGGCAGTGATTGCCATCTGGGGACCTGTCAGGCTTTGTCATTTCCCAGT TTGTTGGTGGTGCCTTTAGTGGTTCCCTAATTTGGGAACACTGATGGGGCCTTGGACAGGGCTTTCTCTC AGGTAGGAGAAATGGGCCCATGATCTCCTCACAGTCGCCCCCAGTCCTTGGCCCTGCTTCCCTGTGTCTC ATGCACTGGCACATATGGTCACCTTGGAGGGCAGACCTAGGAGCCCCTCTGACCACTGAATCCGTCTCCA CACCCCTTCTGCCAAGGGAAGCCCCTTCAGGAAGGACCCCCCAAAGCTGAGGGGCTGAATGTAGCCTTTT CAACAGAGAAGGCTCCCACTTGAGAGCAGCCTCTACCTGACCCCCTGGACCACAGAGAGCCACTCTGACC CTCAGCCCCCTCGCTTCTTCAGCTAAAACTCCAAAGGTTTGGTTTCAGATGGGGTTTGTTTTGTTCTGTT TGGTTTTGGTTTTGTTTGGGGTGGGTGGGTCATTGCGGTCTTAGATTATGTTTCTCTTGCTACCAAACAG TCATGTATTAACTCTCTTTGGATGATGAAGTTTAAAGAGTCAATAAATAGAAACACCAG |
||||||
| Genomic Sequence Description | |||||||
| Genomic Sequence |
|
| Literature |
|---|
| PubMed:11106729 | Kinesin-dependent axonal transport is mediated by the sunday driver (SYD) protein. Bowman AB, Kamal A, Ritchings BW, Philp AV, McGrail M, Gindhart JG, Goldstein LS. Cell. 2000 Nov 10;103(4):583-94. |
| PubMed:8601312 | Interaction of 14-3-3 with signaling proteins is mediated by the recognition of phosphoserine. Muslin AJ, Tanner JW, Allen PM, Shaw AS. Cell. 1996 Mar 22;84(6):889-97 |
| PubMed: 8791419 | The Dbl family of oncogenes. Cerione RA, Zheng Y Curr Opin Cell Biol 1996 Apr;8(2):216-22 |
| PubMed: 10523642 | JSAP1, a novel jun N-terminal protein kinase (JNK)-binding protein that functions as a Scaffold factor in the JNK signaling pathway. Ito M, Yoshioka K, Akechi M, Yamashita S, Takamatsu N, Sugiyama K, Hibi M, Nakabeppu Y, Shiba T, Yamamoto KI Mol Cell Biol 1999 Nov;19(11):7539-48 |
| PubMed:11181572 | Fgd1, the Cdc42 guanine nucleotide exchange factor responsible for faciogenital dysplasia, is localized to the subcortical actin cytoskeleton and Golgi membrane. Estrada L, Caron E, Gorski J. Hum Mol Genet. 2001 Mar 1;10(5):485-95. |
| PubMed:9463375 | In mouse brain profilin I and profilin II associate with regulators of the endocytic pathway and actin assembly. Witke W, Podtelejnikov AV, Di Nardo A, Sutherland JD, Gurniak CB, Dotti C, Mann M EMBO J. 1998 Feb 16;17(4):967-76 |
| Home | Initium | Angiogenesis | FAQ |